A vékonybél anatómiája, élettana és emésztésben betöltött szerepe
A vékonybél anatómiai felépítése
A vékonybél az emésztőcsatorna leghosszabb szakasza, a gyomor és a vastagbél között helyezkedik el. A tápanyagfelszívódás szempontjából a vékonybél az elsőszámú anatómiai struktúra. A vékonybél ezen kívül az emberi szervezet „legnagyobb” immunszerve is. Számos biológiailag aktív amint és pepiidet termel, amelyek részben endokrin, részben parakrin módon szabályozzák az emésztés, motilitás és felszívódás folyamatát.
3 fő szakasz
A vékonybél 3 fő szakasza a duodenum (nyombél, patkóbél), a jejunum (éhbél) és az ileum (csípőbél). A patkóbél elnevezés a bélszakasz alakjára utal. A duodenum szó a hosszúságából ered, a latin duodenum digitorum kifejezés jelentése: 12 ujjnyi. Az éhbél elnevezés arra utal, hogy boncoláskor általában üresen találják, mert perisztaltikája egy ideig a halál beállta után is működik. A csípőbél a hasüreg alsó részéig terjed, jobb és baloldalon a csípőlapátok szomszédságáig.
A vékonybél teljes hossza kb 4-7 méter. Átmérője az orális felén 2,5-3 cm, az aborális felén 2-2,5 cm. A duodenum és a jejunum közötti határ a duodenum lefutása, a környező parenchymalis szervek és érképletek, illetve a Treitz-szalag alapján makroszkóposán jól felismerhető. A jejunum és ileum közötti határ már nem különül el élesen, a szövettani átmenet is fokozatos.
Belső felépítése
A vékonybél-nyálkahártya szervezetünk legnagyobb felülete, ahol a külvilággal érintkezik. Redőzete, az ún. Kerkring-redők makroszkópos szinten szolgálják a felszívódáshoz szükséges minél nagyobb felület kialakítását. A tápanyagok felszívódásának anatómiai egysége a bélboholy (villus), amely a rajta található mikrovillusokkal együtt kb. 200 m2 összfelületet biztosít a felszívódáshoz. A mikrovillusokon helyezkednek el az ún. kefeszegélyenzimek, amelyek az összes tápanyagféleség felszívódásához elengedhetetlen, végső emésztési fázist végzik.
Ezek az enzimek bontják le a diszacharidokat, oligo- és polipetideket és a lipideket. Az emésztésen kívül a vékonybél-nyálkahártyában történik a víz és az elektrolitok felszívódása is, noha ez kitüntetetten a vastagbél feladata. Kizárólag a duodenumban találhatók a Brunner-mirigyek (glandulae duodenales), amelyek a submucosában helyezkednek el, és bőségesen tartalmaznak nyáktermelő kehelysejteket. Ezek a mirigyek a Lieberkühn-cryptákban lévő mirigyekkel együtt termelik a vékonybélnedvet, amely részben az emésztést, részben a nyálkahártya védelmét szolgálja.
- Fitten egy életen át – gyakorlatsorok
- Fitt mindennapok, motiváció a testmozgáshoz
- A rugalmasság növelése edzéssel
Ezek a cikkek is érdekelhetnek:
A vékonybél-nyálkahártyában helyezkednek el azok a sejtes és nem sejtes anatómiai struktúrák is, amelyek a tápcsatorna saját neuroendokrin rendszerét képezik. A vékonybélben található neurokrin-endokrin-parakrin elválasztású mirigyekben termelődik a bioaktív peptidek (gasztrointesztinális hormonok) túlnyomó része.
A lamina propriában és a submucosában elhelyezkedő lymphoid sejtes aggregátumok (pl. a Peyer-plakkok) képezik az intesztinális immunrendszer vázát, amely a szervezet immunglobulint termelő sejtjeinek kb. háromnegyedét tartalmazza. Ezt a rendszert „gut associated lymphoid tissue”-nak (GALT) nevezzük. A GALT-ban termelődő egyik legfontosabb immunglobulin a szekretoros IgA, amely a szervezet első védelmi vonalát képezi az exogén antigénekkel szemben.
A vékonybél a vegetatív idegrendszer szimpatikus beidegzését a truncus sympathicusból, a paraszimpatikus beidegzést a nervus vagustól kapja. A n. vagus pregang-lionaris rostjai a bélfalban érintkeznek az intramuralis plexusokkal, a submucosában a Meissner-féle plexussal, a tunica muscularisban az Auerbach-féle plexussal.
A vékonybél abszorpciós és szekréciós működése
A vékonybél-nyálkahártyán át kétirányú transzport folyik: a bél lumenéből a vér- és nyirokkeringés felé (felszívódás – abszorpció), és a keringésből a lumen felé (kiválasztás – szekréció). A két folyamat közül az abszorpció irányában lényegesen nagyobb a transzportaktivitás. A szervezet működéséhez nélkülözhetetlenül szükséges anyagok membrántranszportja több különböző mechanizmus által valósul meg.
A legegyszerűbb diffúzión kívül további mechanizmusok az ozmózis, illetve a speciális molekuláris (carrier-fehérjék), vagy mikrostrukturális transzferrendszerek (endocytosis) által végzett, energiaigényes aktív transzport. Egyes anyagok nem a nyálkahártya sejtjein át, hanem azok mellett (között) jutnak el a keringésbe (paracellularis út).
A duodenumban keveredik össze az epe és a hasnyálmirigy-nedv a vékonybélnedvvel. Ennek az emésztőnedveket változó összetételben tartalmazó keveréknek hatására indul meg a bevitt táplálék emésztése, a felszívódásra való alkalmassá tétele. Ez a folyamat a duodenumban a legkifejezettebb, a jejunumban az aktivitás már lényegesen kisebb.
A fehérje-, szénhidrát- és zsíremésztés legfontosabb lépcsői a következők:
A fehérjék emésztése és felszívódása
- A gyomorban a pepszin és a sósav hatására megkezdődik a fehérjeemésztés, ami a vékonybélben leáll, mert az itt uralkodó közel neutrális pH a pepszint inaktiválja.
- A hasnyálmirigy-nedvben lévő fehérjeemésztő elő enzimek, a tripszinogén és a chymotripszinogén a duodenumba kerülnek és aktiválódnak.
- A belőlük képződött tripszin és chymotripszin mellett a fehérjék megemésztésében még további két enzimféleség vesz részt. Ezek a szintén a hasnyálmirigyben termelődött karboxipeptidázok és a vékonybél nyálkahártyáján képződött aminopeptidázok, amelyek által a peptidek mindkét terminálisukon hasíthatóvá válnak.
- A fenti enzimek segítségével a fehérjék végül különböző aminosavakból álló kisebb peptidekké bomlanak le.
- Ezeket a kisméretű peptideket a peptidázok végül aminosavakká, di- vagy tripeptidekké bontják le, amelyek a fent leírt különböző mechanizmusok által szívódnak fel.
A szénhidrátok emésztése és felszívódása
A szénhidrátok nagyobb része poliszacharidok (keményítő) formájában kerül a szervezetünkbe. Felszívódásuk monoszacharida formájában történik.
- A poliszacharidok emésztése már a szájüregben megkezdődik a nyálmirigyek alfa-amiláz enzime (ptyalin) által.
- Az így keletkezett poliszacharid fragmentumok a dextrinek.
- A gyomorban a szénhidrátemésztés leáll, mert a savas gyomor nedv a ptyalint inaktiválja.
- A vékonybélben a hasnálmirigy eredetű alfa-amilázok ismét megkezdik a szénhidrátok emésztését.
- Az alfa-amilázok a vékonybél-nyálkahártya sejtjei által termelt glukozidázokkal együtt folytatják a szénhidrátok lebontását, ekkor keletkeznek a maltóz, izomaltóz és glükóz molekulák.
- A vékonybél kefeszegély termeli a maltáz és izomaltáz enzimeket, amelyek hatására a maltózból és az izomaltózból is glükóz képződik, ami a máj felé transzponálódik.
- A szénhidrátok kisebb hányada diszacharid formában (szacharóz és laktóz) kerül a vékonybélbe.
- A szacharózt a kefeszegélyben levő szaharáz fruktózra és glükózra, a laktáz a laktózt (tejcukor) galaktózra és glükózra bontja.
A zsírok emésztése és felszívódása:
- A szervezetbe került zsírok 90%-a triglicerid (neutrális zsír), 10% foszfolipid, koleszterin és zsíroldékony vitaminok (A, D, E és K)
- A trigliceridek hasítása már a gyomor savas közegében megkezdődik a nyálmirigyek által termelt enzim és a gyomorlipázok hatására.
- A zsíremésztés legnagyobb része a vékonybélben zajlik, miután az előemésztett étel az epével és a hasnyálmirigy-nedvvel találkozik.
- A hasnyálmirigy által termelt lipáz a triglicerideket monogliceridekké és szabad zsírsavakká hasítja.
- A koleszterin-zsírsav komplexek és a foszfolipidek feloldása a vékonybél disztális szakaszában megy végbe.
- Az epesavak segítségével a monoglicerideket, zsírsavakat, koleszterint, foszfolipideket és zsíroldékony vitaminokat tartalmazó micellák képződnek.
- Csak a micellák képesek a vékonybél-nyálkahártya mikrovillusaihoz kitapadni és felszívódni.
- A zsírok és építőelemeik felszívódása túlnyomóan a duodenumban és a jejunum kezdeti szakaszán zajlik.
- A rövid és közepesen hosszú szénláncú zsírzsavak diffúzió útján jutnak a bélbolyhok kapillárisaiba, majd onnan a portalis rendszeren át a májba.
- A nagyobb zsírmolekulákat a hámsejtek endocytosissal veszik fel.
- A képződő zsír-fehérje cseppecskék a kilomikronok.
- A kilomikronok a bélbolyhok nyirokkapillárisain át jutnak el a nagyobb nyirokereken keresztül a ductus thoracicusba, majd a keringésbe.
A zsíroldékony vitaminokat a zsírtartalmú táplálékokkal együtt vesszük fel, mivel ezek a vitaminok csak a zsírsavak jelenlétében képződő micellák formájában kerülhetnek reszorpcióra. Ezzel szemben a vízoldékony vitaminok többsége, mint pl. a B- és C-vitaminok, passzívan, diffúzió útján kerülnek felvételre. Speciális a Bi2-vitamin helyzete, ami csak a gyomorban termelődő intrinsic faktorhoz (IF) kötődve tud a terminális ileumból felszívódni.
A vékonybél orális feléből szívódnak fel a szénhidrátok, az aminosavak, a vas, és a zsírok (a szabad zsírsavat és glicerint tartalmazó micellák formájában). A vékonybél alsó feléből jut be szervezetünkbe a koleszterin, a glicerin, az epesavak és B12-vitamin, utóbbi kettő a terminális ileumból.
A vékonybél motilitásának szabályozása
A vékonybél-motilitás modern ismeretei a 20. század elején a béltörvény felfedezésével (Bayliss és Starling) kezdődtek. A tápcsatornában ciklusosán megújuló kontraktilis tevékenységet részletesen Szurszewski jellemezte, és 1969-ben ő nevezte el migráló motoros komplexumnak (MMC) ami jelentős lökést adott a vékonybél-motilitás szabályozásában szerepet játszó mechanizmusok kutatásának is. Az alapkutatások eredményei felkeltették a klinikusok érdeklődését, ami azóta igen jelentős előrehaladást eredményezett a vékonybél-motilitás kórfolyamatainak a megértésében is.
A vékonybél motoros aktivitásának két legfontosabb célja a tápanyagok eljuttatása a felszívódásuk helyére és a salakanyagok továbbítása. A táplálék megemésztése a felszívódás elengedhetetlen feltétele, amihez a táplálék és az emésztőenzimek összekeveredése szükséges, amelyet a vékonybél szegmentáló kontrakciói biztosítanak. Ezen bélmozgások anatómiai alapját a bélfal simaizomzata képezi, amely tartalmazza a kontrakciókat kezdeményező és az aktivitást szabályozó struktúrákat is.
A szabályozásban elsődleges szerepet az intrinsic neurogén, miogén és hormonális mechanizmusok játszanak, de jelentős moduláló hatást gyakorol a központi idegrendszer és a hormonális rendszer, valamint maga a táplálékbevitel is. A táplálékmolekulák érzékelése képezi azoknak a fontos visszacsatolt információknak a forrását, amelyek a motilitást is alapvetően befolyásolják. Mivel az ezen ingerek által kiváltott érzetek szenzációt nem képeznek – megkülönböztetésül a tudatosuló, pl. fájdalomérzetet közvetítő szenzoros idegektől -, afferens innervációnak hívjuk őket. Ebben az értelemben a n. vagus szenzoros rostjainak 80%-a afferens neuron.
A vékonybél-motilitás intrinsic neurogén regulációja – Az enterális idegrendszer (ENS)
A vékonybél-motilitás intrinsic neurogén regulációjának az alapja az enterális idegrendszer (ENS), amely két fő hálózatot alkot: a nyálkahártya és a bélfal körkörös izomrétege között elhelyezkedő submucosus plexust, valamint a tunica muscularis körkörös és hosszanti rétege közötti myentericus plexust. A plexusok idegsejtjei ganglionokba csoportosulnak, amelyeket axonjaik kötegei egymással összekapcsolnak, így alkotnak hálózatokat.
Elhelyezkedésének megfelelően a submucosus plexus a nyálkahártya szekréciós tevékenységének, illetve vérkeringésének szabályozásában játszik elsődleges szerepet. A myentericus plexus fő feladata a vékonybél-motilitás koordinálása. A vékonybél körkörös izomrétegében az izomrostok között egy mély muscularis plexusnak nevezett, sejttestek nélküli idegköteg-hálózat is kimutatható. Funkcionális szempontból a plexusokban lévő neuronok lehetnek primer afferens, motoros és interneuronok.
Az intrinsic primer afferens neuronok sejttestjei a ganglionokban helyezkednek el, egyetlen axonjuk a nyálkahártya epithelsejtjei közelében végződik, és elsősorban kémiai információkat érzékelnek, de vannak falfeszülést érzékelő mechanoszenzoros és nociceptív funkciót ellátó neuronok is. Az interneuronokkal és intrinsic motoneuronokkal képzett szinapszisaik révén alkotják a perisztaltika, a szekréció és a mucosa vérkeringését szabályozó intrinsic reflexeket, amelyeket a denervált bélen is megfigyelhetünk.
Az ingerületet az interneuronok legfeljebb néhány centiméterre közvetítik. Az ennél nagyobb reflexívek az intesztinofugális afferens neuronok közreműködésével jönnek létre, amelyeknek a sejttestjei szintén a bélfalban helyezkednek el, de a prevertebralis ganglionokban zajló szinaptikus információátvitel révén jöhetnek létre a bél távolabbi részei közötti reflexes folyamatok. Ezek a reflexek elsősorban entero-enteralis inhibitoros folyamatokat szabályoznak.
A motoros neuronok axonjai beidegzik a hosszanti és körkörös izomrétegeket, amelyek szimultán kontrakciója eredményezi a propulzív kontrakciókat. A serkentő motoros beidegzés acetilkolin és substance P, a gátló hatás pedig nitrogén-monoxid (NO), vazoaktív intesztinális peptid (VIP) és ATP-traszmitterek közvetítésével történik. Az NO-tartamú neuronok jelentős része kalcitonin génkapcsolt pepiidet (CGRP) is termel, ami szintén simaizom-relaxációt okoz.
A simaizom kontraktilis tevékenységét alapvetően a Cajal-sejtek (ICC) által generált elektromos aktivitás koordinálja. Az ICC-k generálják a lassú hullámokat, amelyek a vékonybél-kontrakciók ritmusának az alapját képezik. A simaizom-syncytiumok, a neuronok és ICC-k egymással gap junctiókkal kapcsolódva hozzák létre a kontraktilis egységet. Az izomsejtekben intracelluláris kalcium-felszabadulás aktiválja az aktin- és miozin-filamentumokat, és kontrakció következik be.
A vékonybél extrinsic beidegzése
A vékonybél extrinsic beidegzését kizárólag a nervus vagus paraszimpatikus és thoracicus szimpatikus preganglionalis rostjai biztosítják. A paraszimpatikus idegek sejttestjei a vagus dorsalis motoros magjából indulnak ki. A bolygóideg hátsó kötegében, a plexus coeliacusban és az arteria mesenterica superior elágazódásai mentén futó ágaik a gangliononjait elérve elágazódnak, és varicosusan kitágult végződéseikkel szinapszisokat képeznek a serkentő és gátló neuronokkal. így tehát a vagushatás a vékonybél-motilitásra egyaránt lehet serkentő és gátló.
A felszabaduló acetilkolin az enterális idegeken a posztszinaptikus nikotinos receptorokhoz kötődik. A szimpatikus preganglionalis beidegzés a thoracalis 9-12 szegmentumokból a ganglion coeliacumban és a mesenetericum superiusban kapcsolódik át a postganglionalis idegekre, amelyek az arteria coeliaca és az arteria mesenterica superior mentén jutnak el a vékonybélbe. A szimpatikus hatásra felszabaduló noradrenalin gátolja a vékonybél-motilitást, ami egyrészt a kolinerg transzmisszió gátlása, másrészt pedig a simaizomzat a-adrenerg receptorainak aktiválása révén valósul meg.
A vékonybél-motilitás hormonális szabályozása
A vékonybél-motilitás szabályozásában számos hormon is közreműködik. A motilin és ghrelin az MMC kiváltásában, a szomatosztatin és az enkefalinok pedig vékonybél mentén végbemenő propagációjában játszik szerepet. A kolecisztokinin (CCK), inzulin, gasztrin, neurotenzin, leptin digesztív fázist vált ki, viszont a hypothalamus ventromedialis területén felszabaduló CCK az interdigesztív állapotot tartja fenn, az orexin-A pedig megnyújtja az MMC-ciklusok hosszát. A kórfolyamatokban a corticotropin releasing factor (CRF), thyrotropin releasing hormon (TRH) és számos citokin (IL-1(3, TNFa) is fontos szerepet tölt be.
A perisztaltikus tevékenység és az MMC-aktivitás elektrofiziológiai alapjai
Elektromiográflás vizsgálatokkal a vékonybélben folyamatos, ciklikus, oszcilláló elektromos aktivitás regisztrálható, amelyeket lassú hullámoknak, vagy pacemakerpotenciálnak nevezünk. Nem minden lassú hullám vált ki a simaizomban kontrakciót. A lassú hullámok frekvenciája meghatározza az izomkontrakciók maximális számát. Elektromiográflás felvételeken a kontrakciókat a lassú hullámokon megjelenő tüskesorozatok (spike bursts) mutatják.
Alaphelyzetben neuromuscularis junkciókban a kontrakciót a gátló hatású motoneuronokból folyamatosan felszabaduló neurotranszmitterek megakadályozzák. Fázikus kontrakció akkor következik be, amikor az interneuronok inaktiválják az inhibitoros motoneuronokat, és így felszabadítják a simaizomsejteket a gátlás alól. Tehát a gátló motoneuronok nettó aktivitása határozza meg, hogy a folyamatosan jelenlévő lassú hullám hol és mikor válthat ki kontrakciót, továbbá azt is, hogy a kontrakció merre és meddig terjedhet.
Az afferens neuronok által érzékelt inger keletkezési helyétől orális irányban fokozza a motoros aktivitást, kontrakciókat vált ki (felszálló serkentő hatás), míg aborális irányban gátlást eredményez, és az izomzat ellazul (leszálló gátló hatás). E két motilitási alapjelenség térben és időben összerendezett működése felelős a béltartalom előre vagy retrográd irányú mozgatásáért. A perisztaltikus kontrakciók a bélben manometriával mérhető nyomással és a béltartalom tovahaladásával, tehát a tranzitidővel jellemezhetők.
Ezeknek a paramétereknek az intenzitása, térbeli és időbeli lezajlása a posztprandiális és étkezések közötti (interdigesztív) időszakokban különböző, amelyeket elsődlegesen a lumenben jelenlévő táplálék által kiváltott inger tulajdonságai határoznak meg. Az étkezés alatt és közvetlenül azt követően számos kémiai és fizikai inger éri a belet, amelyek a bélben intenzív, gyors ritmusban ismétlődő, fázikus kontrakti-intracelluláris elektromos aktivitást indukálnak, amelyek fiziológiás célja a táplálék továbbítása és szétterjesztése a bél felszívó felületén.
A táplálék és nyálkahártya közötti direkt kontaktus elengedhetetlen feltétele a digesztív motoros tevékenység kiváltásának, de hozzájárulnak a felszabaduló enterohormonok (CCK, GLP-1) is. A digesztív perisztaltika időtartama és intenzitása függ a táplálék mennyiségétől, fehérje-, szénhidrát-, zsír- és energiatartalmától. A nem felszívódó béltartalom csökkenti a vékonybélben a tranzitidőt. Étkezés után 4-6 órával, amikor már jelentős béltartalom a bélben nincs jelen, a bélben detektálható éhomi (interdigesztív) motoros tevékenységet nevezzük migráló motoros komplexumnak (MMC).
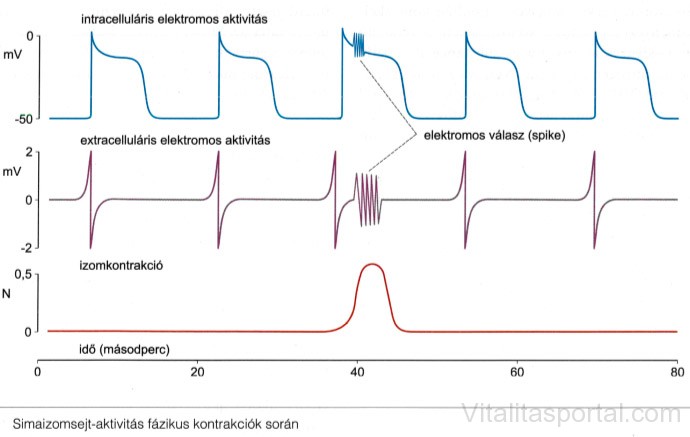
Manometriás vagy elektromiográfiás mérésekkel az interdigesztív motoros aktivitásnak három fázisát különböztetjük meg.
Az I. fázis könnyen felismerhető, a nyugalom időszaka, amikor a kontrakciók száma 10 perc alatt kevesebb, mint három. A harmadik fázis szintén könnyen felismerhető a magas amplitúdójú, az adott bélszakaszra jellemző maximális frekvenciájú kontraktilis aktivitásról. Az első és harmadik fázis közötti II. fázist a nyugalmi fázisnál magasabb, de a régióra jellemző maximális frekvenciájú kontraktilis tevékenységnél alacsonyabb frekvenciájú, alacsonyabb amplitúdójú nyomáshullámok jellemzik. Az MMC proximálisan kezdődik és disztális irányba haladva különböző távolságokra jut el, de csak kevés III. fázis éri el az ileumot.
Továbbá, a III. fázis, különböző helyeken kezdődhet; csak mintegy harmad részük gastroduodenalis kiindulású, a többségük a jejunum proximális szakaszán kezdődik. Az MMC időtartama emberben 80-120 perc, de jelentős interindividuális variabilitás jellemző. Figyelembe véve a vékonybél hosszát, nyilvánvaló, hogy egy adott időpontban a vékonybél egy része az MMC első, míg másik része a második, vagy harmadik fázisában van. Az MMC fiziológiás célja a bél tisztántartása, tehát a nem felszívódó anyagok és vékonybélbe jutó baktériumok továbbítása a vastagbélbe, ami túlnyomórészt a III. fázis során következik be.

Szerző: Griffel Tibor
Végzettség: ELTE – Eötvös Loránd Tudományegyetem. Szakterület: a szív- és érrendszeri betegségek, gasztroenterológiai betegségek és a légzőrendszeri betegségek. Jelenleg reflexológus, életmód és tanácsadó terapeuta tanulmányokat is végzek.